
Note: the original web site describing our primer sequences expired in July 2017; I have reposted the original information here. Click here for a pdf version. -R. Vilgalys, Aug 2018
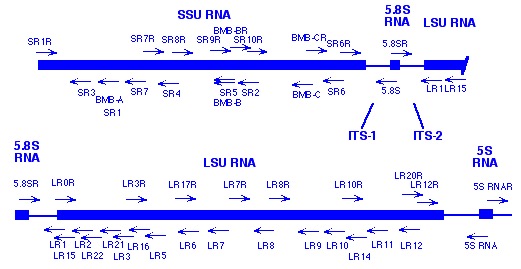
Primer sites within the ribosomal RNA genes of fungi
Over the years, our lab has compiled a useful list of conserved primer sequences useful for amplification and sequencing of nuclear rDNA from most major groups of fungi (primarily Eumycota), as well as other eukaryotes. All of these primers were first identified and tested by our own lab based on consensus between the published large and small subunit RNA sequences from fungi, plants and other eukaryotes; sources of other useful primer sequences from published literature are also indicated. Together, these primers span most of the nuclear rDNA coding region (see figures), permitting amplification of any desired region. Standard symbols are used for the four primary nucleotides; variable positions are indicated as follows: P=A,G / Q=C,T / R=A,T / V=A,C / W=G,T. Primers ending with “R” represent the coding strand (same as RNA). All other primers are complementary to the coding strand. This information is provided freely and may be passed on to anyone who wants to use it.
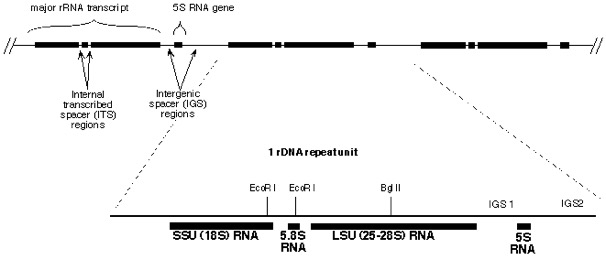
Organization of ribosomal RNA genes in fungi. Multiple copies of near-identical rDNA operons are organized in a head-to-toe manner on a single chromosome.
The nuclear-encoded ribosomal RNA genes (rDNA) of fungi exist as a multiple-copy gene family comprised of highly similar DNA sequences (typically from 8-12 kb each) arranged in a head-to-toe manner. Each repeat unit has coding regions for one major transcript (containing the primary rRNAs for a single ribosome), punctuated by one or more intergenic spacer (IGS) regions. In some groups (mostly basidiomcyetes and some ascomycetous yeasts), each repeat also has a separately transcribed coding region for 5S RNA whose position and direction of transcription may vay among groups. Several restriction sites for EcoRI and BglII are conserved in the rDNA of fungi. Nearly all basidiomycetes we’ve studied share an EcoRI site within the 5.8S RNA gene along with a BglII site halfway into the LSU RNA sequence. Primers 5.8SR and LR7 include these restriction sites, which makes them convenient for cloning.
For those who aren’t familiar with rDNA and fungal systematics, several excellent reviews are available on fungi (Hibbett, 1992) and generally for eukaryotes (Hillis and Dixon, 1991). See Gerbi (1986) for a general introduction to the molecular biology and evolution of rDNA in other eukaryotes. Another useful source of primer information may be found in Gargas & Depriest (1996) and at the Tom Bruns lab web site http://mendel.berkeley.edu/boletus/boletus.html.
| SSU (small-subunit 18S RNA) primer sequences | ||
| Primer name | Sequence (5′–>3′) | Position within S. cereviseae 17S RNA |
| BMB-‘A’ | GRATTACCGCGGCWGCTG | 580-558 |
| BMB-‘B’ | CCGTCAATTCVTTTPAGTTT | 1146-1127 |
| BMB-‘C’ | ACGGGCGGTGTGTPC | 1638-1624 |
| BMB-BR | CTTAAAGGAATTGACGGAA | 1130-1148 |
| BMB-CR | GTACACACCGCCCGTCG | 1624-1640 |
| SR1R | TACCTGGTTGATQCTGCCAGT | 564-578 |
| SR1 | ATTACCGCGGCTGCT | 578-564 |
| SR2 | CGGCCATGCACCACC | 1277-1263 |
| SR3 | GAAAGTTGATAGGGCT | 318-302 |
| SR4 | AAACCAACAAAATAGAA | 838-820 |
| SR5 | GTGCCCTTCCGTCAATT | 1146-1130 |
| SR6 | TGTTACGACTTTTACTT | 1760-1744 |
| SR6R | AAGWAAAAGTCGTAACAAGG | 1744-1763 |
| SR7 | GTTCAACTACGAGCTTTTTAA | 617-637 |
| SR7R | AGTTAAAAAGCTCGTAGTTG | 637-617 |
| SR8R | GAACCAGGACTTTTACCTT | 732-749 |
| SR9R | QAGAGGTGAAATTCT | 896-910 |
| SR10R | TTTGACTCAACACGGG | 1181-1196 |
| NS1 | GTAGTCATATGCTTGTCTC | |
| NS2 | GGCTGCTGGCACCAGACTTGC | |
| NS3 | GCAAGTCTGGTGCCAGCAGCC | |
| NS4 | CTTCCGTCAATTCCTTTAAG | (similar to BMB-B) |
| NS5 | AACTTAAAGGAATTGACGGAAG | (is similar to BMB-BR) |
| NS6 | GCATCACAGACCTGTTATTGCCTC | |
| NS7 | GAGGCAATAACAGGTCTGTGATGC | |
| NS8 | TCCGCAGGTTCACCTACGGA | |
| BMB = “universal” SSU primers developed by Lane et al., 1985 | ||
| SR = primers developed by Vilgalys lab | ||
| NS = primers described by White et al., 1990 | ||
| LSU (large subunit 25-28S RNA) | |||
| Primer name | Sequence (5′–>3′) | Position within S. cereviseae rRNA | comments |
| 5.8S | CGCTGCGTTCTTCATCG | 51-35 (5.8S RNA) | contains EcoRI site |
| 5.8SR | TCGATGAAGAACGCAGCG | 34-51 (5.8S RNA) | contains EcoRI site |
| LR0R | ACCCGCTGAACTTAAGC | 26-42 | |
| LR1 | GGTTGGTTTCTTTTCCT | 73-57 | |
| LR2 | TTTTCAAAGTTCTTTTC | 385-370 | |
| LR2R | AAGAACTTTGAAAAGAG | 374-389 | |
| LR3 | CCGTGTTTCAAGACGGG | 651-635 | |
| LR3R | GTCTTGAAACACGGACC | 638-654 | |
| LR4 | ACCAGAGTTTCCTCTGG | 854-838 | |
| LR5 | TCCTGAGGGAAACTTCG | 964-948 | |
| LR6 | CGCCAGTTCTGCTTACC | 1141-1125 | |
| LR7 | TACTACCACCAAGATCT | 1448-1432 | contains BglII site |
| LR7R | GCAGATCTTGGTGGTAG | 1430-1446 | contains BglII site |
| LR8 | CACCTTGGAGACCTGCT | 1861-1845 | |
| LR8R | AGCAGGTCTCCAAGGTG | 1845-1861 | |
| LR9 | AGAGCACTGGGCAGAAA | 2204-2188 | |
| LR10 | AGTCAAGCTCAACAGGG | 2420-2404 | |
| LR10R | GACCCTGTTGAGCTTGA | 2402-2418 | |
| LR11 | GCCAGTTATCCCTGTGGTAA | 2821-2802 | |
| LR12 | GACTTAGAGGCGTTCAG | 3124-3106 | |
| LR12R | CTGAACGCCTCTAAGTCAGAA | 3106-3126 | |
| LR13 | CGTAACAACAAGGCTACT | 3357-3340 | |
| LR14 | AGCCAAACTCCCCACCTG | 2616-2599 | |
| LR15 | TAAATTACAACTCGGAC | 154-138 | |
| LR16 | TTCCACCCAAACACTCG | 1081-1065 | |
| LR17R | TAACCTATTCTCAAACTT | 1033-1050 | |
| LR20R | GTGAGACAGGTTAGTTTTACCCT | 2959-2982 | |
| LR21 | ACTTCAAGCGTTTCCCTTT | 424-393 | |
| LR22 | CCTCACGGTACTTGTTCGCT | 364-344 | |
Note: Many of these primers were first reported in several papers (Hopple and Vilgalys, Hibbett and Vilgalys, Vilgalys and Hester, ). Most molecular systematics studies only utilize the first 600-900 bases from the LSU gene, which includes three divergent domains (D1, D2, D3) that are among the most variable regions within the entire gene (much of the LSU is invariant even across widely divergent taxa). Most of the data in our Agaricales LSU database consists of the first 900 bases from the LSU gene (we typically amplify using primers 5.8SR + LR7, followed by sequencing using primers LR5, LR16, LR0R, and LR3R).
| ITS (internal transcribed spacer) region primers | |||
| primer name | sequence (5′->3′) | comments | reference |
| ITS1 | TCCGTAGGTGAACCTGCGG | White et al, 1990 | |
| ITS2 | GCTGCGTTCTTCATCGATGC | (similar to 5.8S | White et al, 1990 |
| ITS3 | GCATCGATGAAGAACGCAGC | (similar to 5.8SR) | White et al, 1990 |
| ITS4 | TCCTCCGCTTATTGATATGC | White et al, 1990 | |
| ITS5 | GGAAGTAAAAGTCGTAACAAGG | (similar to SR6R) | White et al, 1990 |
| ITS1-F | CTTGGTCATTTAGAGGAAGTAA | Gardes & Bruns, 1993 | |
| ITS4-B | CAGGAGACTTGTACACGGTCCAG | Gardes & Bruns, 1993 | |
| 5.8S | CGCTGCGTTCTTCATCG | Vilgalys lab | |
| 5.8SR | TCGATGAAGAACGCAGCG | Vilgalys lab | |
| SR6R | AAGWAAAAGTCGTAACAAGG | Vilgalys lab | |
Notes on ITS: The ITS region is the most widely sequenced DNA region for Fungi. It has typically been most useful for molecular systematics at the species level, and even within species (e.g., to identify geographic races). Because of its higher degree of variation than other genic regions of rDNA (SSU and LSU), variation among individual rDNA repeats can sometimes be observed within both the ITS and IGS regions. In addition to the standard ITS1+ITS4 primers used by most labs, numerous taxon-specific primers have been described that allow selective amplification of fungal sequences (e.g., see Gardes & Bruns 1993 paper describing amplification of basidiomycete ITS sequences from mycorrhiza samples).
| IGS (intergenic spacer primers, including 5S RNA primer sequences primarily for basidiomycete fungi) | |||
| primer name | sequence (5′->3′) | comments | reference |
| LR12R | GAACGCCTCTAAGTCAGAATCC | located within the LSU RNA (see above) | Vilgalys lab |
| invSR1R | ACTGGCAGAATCAACCAGGTA | located within the SSU RNA (positions 21-1) | Vilgalys lab |
| 5SRNA | ATCAGACGGGATGCGGT | (complementary to 5S RNA positions 46-26) | Vilgalys lab |
| 5SRNAR | ACQGCATCCCGTCTGAT | (5S RNA positions 26-46) | Vilgalys lab |
Notes on IGS region: The greatest amount sequence variation in rDNA exists within the IGS region (sometimes also known as the non-transcribed spacer or NTS region). The size of the IGS region may vary from 2 kb upwards. It is not unusual to find hypervariability for this region (necessitating cloning of individual repeat haplotypes). Several patterns of organization can be found in different groups of fungi: 1. Most filamentous ascomycetes have a single uninterrupted IGS region (between the end of the LSU and start of the next SSU sequence), which may vary in length from 2-5 kb or more. Amplification of the entire IGS region requires using primers anchored in the 3′ end of the LSU gene (e.g., LR12R) and 5′ end of the SSU RNA gene (e.g., invSR1R); 2. In many ascomycetous yeasts and nearly all basidiomycetes, the IGS also contains a single coding region for the 5S RNA gene, which divides the IGS into two smaller regions that may be more easily amplified using. Depending on the orientation and position of the 5S RNA gene, PCR may be used to sequentially amplify a portion of the intergenic spacer region (IGS) beyond the large subunit RNA coding region.
REFERENCES
Bruns, T. D., R. Vilgalys, S. M. Barns, D. Gonzalez, D. S. Hibbett, D. J. Lane, L. Simon, S. Stickel, T. M. Szaro, W. G. Weisburg, and M. L. Sogin. 1992. Evolutionary relationships within the fungi: analyses of nuclear small subunit rRNA sequences. Molec. Phylog. Evol. 1: 231-241.
Bruns, T. D., T. J. White, and J. W. Taylor. 1991. Fungal molecular systematics. Ann. Rev. Ecol. Syst. 22: 525-564.
DePriest, P. T., and M. D. Been. 1992. Numerous group I introns with variable distributions in the ribosomal DNA of a lichen fungus. J. Mol. Biol. 228: 315-321.
Elwood, H. J., G. J. Olsen, and M. L. Sogin. 1985. The small subunit ribosomal RNA gene sequences from the hypotrichous ciliates Oxytricha nova and Stylonychia pustula. Mol. Biol. Evol. 2: 399-410.
Gardes, M., and T. D. Bruns. 1993. ITS primers with enhanced specificity for basidiomycetes – application to the identification of mycorrhizae and rusts. Mol. Ecol. 2: 113-118.
Gargas, A., and P.T. DePriest. 1996. A nomenclature for fungal PCR primers with examples from intron-containing SSU rDNA.
Mycologia 88: 745-748
Gargas, A., and J.W. Taylor. 1992. Polymerase chain reaction (PCR) primers for amplifying and sequencing 18S rDNA from
lichenized fungi. Mycologia 84: 589-592.
Gerbi, S. A. 1986. Chapter 7 – Evolution of ribosomal DNA. Pp. 419-517 In: Molecular evolution, ed. McIntyre, R.
Hibbett, D. S. 1991. Phylogenetic relationships of the Basidiomycete genus Lentinus: evidence from ribosomal RNA and morphology. Ph.D. Thesis, Duke University, 1991.
Hibbett, D. S. 1992. Ribosomal RNA and fungal systematics. Trans. Mycol. Soc. Jpn. 33: 533-556.
Hibbett, D. S., and R. Vilgalys. 1991. Evolutionary relationships of Lentinus to the Polyporaceae: evidence from restriction analysis of enzymatically amplified ribosomal DNA. Mycologia 83: 425-439.
Hibbett, D. S., and R. Vilgalys. 1993. Phylogenetic relationships of the Basidiomycete genus Lentinus inferred from molecular and morphological characters. Syst. Bot. 18: 409-433.
Hillis, D. M., and M. T. Dixon. 1991. Ribosomal DNA: molecular evoluiton and phylogenetic inference. Quart. Rev. Biol. 66: 411-453.
Hopple JS, Vilgalys R (1999) Phylogenetic relationships in the mushroom genus Coprinus and dark-spored allies based on sequence data from the nuclear gene coding for the large ribosomal subunit RNA: divergent domains, outgroups, and monophyly. Molecular Phylogenetics and Evolution 13:1–19. doi: 10.1006/mpev.1999.0634
Hopple, J. S., Jr., and R. Vilgalys. 1994. Phylogenetic relationship among coprinoid taxa and allies based on data from restriction site mapping of nuclear rDNA. Mycologia 86: 96-107.
Lane, D. J., B. Pace, G. J. Olsen, D. A. Stahl, M. L. Sogin, and N. R. Pace. 1985. Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc. Natl. Acad. Sci., U. S. A. 82: 6955-6959.
Vilgalys, R., and D. Gonzalez. 1990. Organization of ribosomal DNA in the basidiomycete Thanatephorus praticola. Curr. Genet. 18: 277-280.
Vilgalys R, Hester M (1990) Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J Bacteriol 172:4238–4246.
Vilgalys, R., J. S. Hopple, Jr., and D. S. Hibbett. 1994. Phylogenetic implications of generic concepts in fungal taxonomy: The impact of molecular systematic studies. Mycologica Helvetica 6: 73-91.
White, T. J., T. Bruns, S. Lee, and J. W. Taylor. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. Pp. 315-322 In: PCR Protocols: A Guide to Methods and Applications, eds. Innis, M. A., D. H. Gelfand, J. J. Sninsky, and T. J. White. Academic Press, Inc., New York.