Structural Basis of Cellular Nucleoside Transport
Being that nucleosides are hydrophilic molecules, they require specific membrane transporter proteins—known as nucleoside transporters (NTs)—to cross the cell membrane. Such a process is mediated by two distinct protein families: concentrative and equilibrative nucleoside transporters (CNTs and ENTs). Since nucleoside transport is associated with essential physiological processes, such as nucleic acid synthesis and the termination of adenosine signaling, we think it is important to interrogate the transport mechanisms of NTs in order to gain a deeper understanding of nucleoside-related physiological and drug pharmacology. But the biggest hurdle in revealing these mechanisms had been the lack of atomic structures that reveal the design principles behind CNTs and ENTs.
Solving the Crystal Structures
In the Lee Lab, we solved the crystal structure of a concentrative nucleoside transporter from Vibrio cholerae (vcCNT), the first reported structure of any CNT family protein (Nature 2012). We also determined crystal structures of vcCNT in complex with nucleoside-derived anticancer and antiviral drugs and studied their interactions using functional assays employing ITC, fluorescence-based equilibrium binding, and a radioactive flux assay to understand the principles of substrate recognition by CNTs (eLife 2014). In 2017, we reported the structures of inward-facing, intermediate, and outward-facing states of CNT from Neisseria wadsworthii (Nature 2017).
Our group also solved crystal structures of an equilibrative nucleoside transporter from H. sapiens (human ENT1) the first reported structure of any ENT family protein (Nat Struc Mol Biol 2019). These structures feature human ENT1 in complex with two different adenosine reuptake inhibitors, which are pharmacological agents used to stimulate the tissue-protective effects of adenosine receptor signaling by ENT block.
Our Discoveries
Our research on CNTs provides proof of principle for utilizing transporter structural and functional information to help design compounds so they can enter cells more efficiently and selectively. Taking what we learned in our structural studies, we rationally modified an existing anti-cancer drug, resulting in higher transport selectivity and efficiency through a single human CNT subtype (eLife 2014). Furthermore, CNT is known to operate via an elevator-type transport mechanism, but the structural features concerning the conformational path that leads to such a transition were not yet known. By solving structures of CNT from inward to intermediate- and outward-facing states, we showed direct evidence for the trajectory of conformational transition in the elevator model for the first time (Nature 2017).
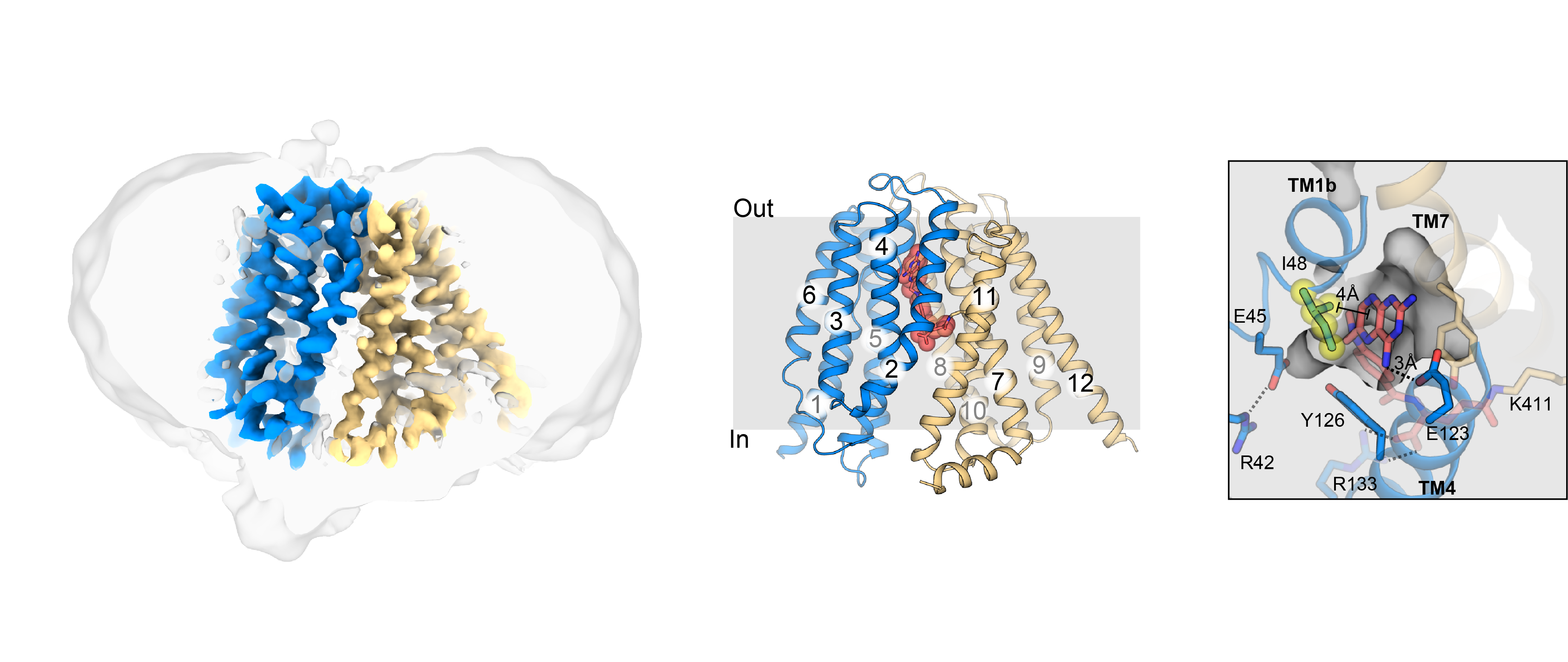
Snapshots of how nucleoside-derived anticancer and antiviral drugs and nucleosides are transported into the cell by concentrative nucleoside transporters (CNTs). The transport domain (yellow) of CNT moves relative to the scaffold domain (blue) with a large elevator-type motion to transport drugs across the cell membrane.
Our work on hENT1 revealed the molecular architecture of the ENT family protein fold, and design principles of human ENT1 inhibition by two different adenosine reuptake inhibitors, while providing initial insight into the nucleoside transport mechanism of ENTs. We also discovered how the reuptake inhibitor NBMPR inhibits this transporter in a unique manner, which sets the stage for the rational design of improved small-molecule therapeutics targeting human ENTs.
Atomic structures of human ENT1 in complex with adenosine reuptake inhibitors. We found that the inhibitors dilazep and NBMPR exhibit distinct structural mechanisms of inhibition.
Antifolate Drug Transport
Folates (vitamin B9) are small anionic biomolecules with widespread roles in cellular metabolism. Being necessary for amino acid metabolism and nucleic acid biosynthesis, rapidly dividing cancer cells are particularly dependent upon this nutrient, as is the immune system. Owing to their charged nature, they rely on transporters and/or internalizing receptors to cross the cell membrane. Our group is interested in folate uptake and efflux mediated by the human reduced folate carrier (hRFC)—one of the most important folate transporters in humans. Due to chemical similarities with dietary and endogenous folates, antifolate therapeutics (such as methotrexate) also rely on hRFC to gain cellular entry to carry out their modes of action. Seeing that hRFC is an important determinant of antifolate drug action, we are motivated to interrogate antifolate drug recognition and transport by hRFC. A roadblock toward this goal had been the lack of an experimental structure of hRFC in complex with an antifolate therapeutic.
Solving the Structure
Since hRFC is a small membrane protein (~60 kDa), structure determination by cryo-EM is difficult. Furthermore, we found methotrexate does not stably bind to detergent-purified hRFC. Despite these challenges, we were successful in solving the cryo-EM structure of hRFC with methotrexate trapped in its central cavity (Nature 2022). We then utilized radiotracer uptake assays and molecular dynamics simulations to further probe how this transporter selectively recognizes folates and antifolate drugs.
Our Discovery
Our study paves the way for a comprehensive understanding of drug recognition and selectivity by hRFC. In addition to revealing detailed features of drug (methotrexate) binding, our study also gained insights into the general principles governing how other antifolates (such as PT523) may interact with this transporter, and how certain natural mutations give rise to drug resistance. Overall, a better understanding of how to more selectively tune drug interaction with hRFC could enable the development of improved therapeutics, and further personalized medicine.
Cryo-EM structure of the human reduced folate carrier with methotrexate trapped in its central cavity. This work uncovers detailed features of drug recognition by hRFC in the context of its inward-facing conformational state.